ELECTROCOCHLEOGRAPHIE
(EcoG)
HISTORIQUE : (2, 17)
En 1926, Adrian fut le premier à enregistrer les potentiels d’action des nerfs sensitifs chez l’animal. En 1930, Wever et Bray enregistrèrent, chez l’animal, les potentiels d’action du nerf cochléaire. Leurs travaux conduisirent à la découverte du phénomène microphonique cochléaire. En 1931 Adrian et coll. présentèrent une étude détaillée du phénomène microphonique chez l’animal. L’intérêt de ce phénomène chez l’homme fut remarqué et motiva d’autres études chez l’animal et chez l’homme. En 1935 (Fromm et coll.) puis en 1939 (Andreev et coll.) et en 1941 (Perlmann et Case), des tentatives d’observation des réponses microphoniques ont été présentées. Bien sûr, les difficultés techniques à l’époque étaient considérables et les résultats incertains.
Plus tard Lempert et coll., en 1947, réalisèrent une nouvelle tentative d’enregistrement du microphonique cochléaire, chez l’homme, en plaçant une électrode sur la membrane de la fenêtre ronde, lors d’une intervention chirurgicale. Ils purent obtenir des résultats satisfaisants, bien que l’amplitude des signaux recueillis fut très faible. Ils proposèrent, ensuite, dans le but de trouver une méthode applicable en dehors de toute chirurgie, la mise en place d’une fine électrode rigide directement sur le promontoire à travers le tympan.
Vers la fin des années 1950, les possibilités techniques s’étant considérablement développées, en même temps que se précisait la nature des réponses cochléaires chez l’animal, ces observations ont été reprises par Ruben et ses collaborateurs du Johns Hopkins Hospital à Baltimore. A la même époque se développait une méthode d’audiométrie objective {potentiels évoqués auditifs (PEA)}, basée également sur l’observation des réponses électriques.
De 1959 à 1967, Ruben a pu présenter des enregistrements extrêmement précis des potentiels cochléaires, en particulier du potentiel d’action global du nerf auditif pour la première fois chez l’homme, que ce soit chez l’adulte ou chez l’enfant et dans diverses situations pathologiques.
Le problème principal était d’extraire et surtout de reconnaître la réponse cochléaire. C’est à dire de vérifier que le signal observé était bien provoqué par la stimulation sonore. L’introduction des moyens électroniques, offrant la possibilité de réaliser une sommation et une moyenne des signaux recueillis, résolut ce problème et permit d’extraire les réponses cochléaires du bruit de fond dans lequel ils étaient noyés.
C’est ainsi qu’en 1967, plusieurs auteurs [ Portmann, Le Bert, Aran, Yoshie, Ohashi, Suzuki, Sohmer, Freinmesser, Spreng, Keidel ], purent publier, quasi simultanément, des études décrivant des différents techniques pour la réalisation de l’EcoG ainsi que ses intérêts cliniques.
Aran et col. introduisirent à partir de 1968 dans leur pratique clinique la réalisation de l’électrocochléographie, en utilisant une électrode rigide posée sur le promontoire à travers le tympan et sous anesthésie générale.
En 1978, Gibson (16) décrivit les principes électrophysiologiques ainsi que les applications cliniques de l’électrocochléographie.
DEFINITION : (2, 3, 17)
L’électrocochléographie (EcoG) est un examen électrophysiologique qui permet l’enregistrement des potentiels électriques de l’organe de Corti et des fibres nerveuses constituant le nerf auditif à l’intérieur de la cochlée, en réponse à une stimulation sonore.
L’activité électrique des structures cochléaires est complexe et difficile à observer au niveau de la cellule sensorielle ou nerveuse. Cependant, elle se manifeste par des phénomènes électriques globaux qui sont une image du fonctionnement de l’oreille interne. Ces phénomènes électriques sont enregistrés par l’EcoG grâce à une électrode qui doit être posée le plus proche possible de la cochlée. Comme nous le verrons plus loin, l’emplacement idéal, chez l’homme, est au niveau de la membrane de la fenêtre ronde.
Les principes généraux de l’EcoG sont similaires aux potentiels évoqués auditifs (PEA).
REPONSES COCHLEAIRES : (2, 17)
Les phénomènes électriques pris en considération sont ceux qui sont provoqués par une stimulation sonore. Sont exclus, donc, les potentiels de repos de la cochlée en l’absence de toute stimulation (potentiels endo-cochléaires).
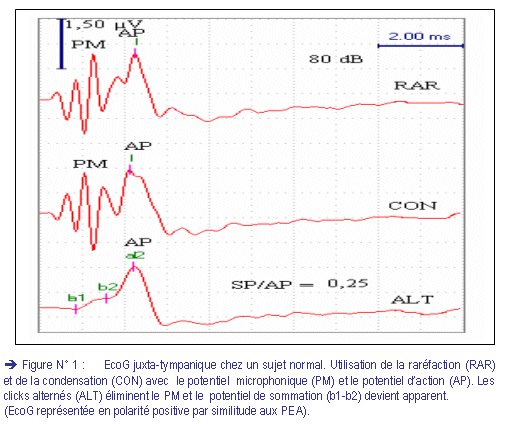
Ces réponses recueillies sont de deux types : (fig. 1)
1) Des réponses sensorielles, dont la source est les cellules ciliées de l’organe de Corti.
2) Des réponses nerveuses, provenant des fibres nerveuses à l’intérieur de la cochlée qui donnent naissance au nerf auditif.
Réponses sensorielles :
Les réponses électriques des cellules ciliées, en réponse à un stimulus sonore, se décomposent en deux potentiels de nature différente.
I) Le potentiel microphonique cochléaire : {Cochlear microphonic (CM)}
C’est un signal électrique, produit par les vibrations des cellules ciliées de l’organe de Corti, qui reproduit exactement, dans certaines limites qui sont celles du champ auditif, la forme de la vibration sonore (en particulier sans aucun délai), comme le ferait un microphone.
Le potentiel microphonique, ne présentant pas de seuil, est de faible amplitude et dépendant des conditions d’enregistrement. Il peut être très aisément confondu avec tout autre phénomène microphonique non cochléaire, provenant par exemple de la vibration de l’électrode.
Découvert depuis très longtemps chez l’animal (phénomène de Wever et Bray, 1930), de part ses propriétés et sa provenance, il ne présente pas d’intérêt clinique et ne semble pas pouvoir servir de base valable à un test électrophysiologique.
Son élimination, lors de l’enregistrement de l’EcoG, est obtenue par l’utilisation des stimuli sonores de phases alternées (Clics alternés). En effet, à l’inverse des autres réponses cochléaires, lors de deux stimulations sonores identiques, mais de phases opposées, le potentiel microphonique d’une stimulation à l’autre s’inverse de la même manière. En additionnant dans la mémoire du moyenneur les réponses successives à deux séries de stimulations l’une dite positive (polarité de l’impulsion servant à produire le clic), l’autre de phase opposée (inversion de la polarité de l’impulsion), on obtient en même temps non seulement l’élimination du bruit du fond aléatoire et l’augmentation de l’amplitude de la réponse nerveuse qui s’additionne, mais encore l’élimination de la réponse microphonique. Puisque la moyenne des signaux de phases opposées est nulle (fig. 1).
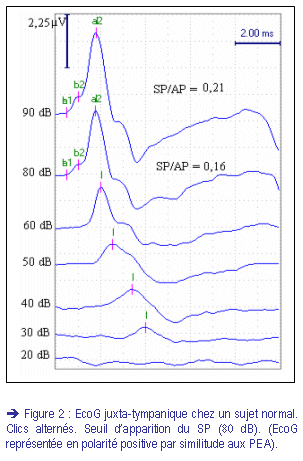
II) Le potentiel de sommation : {Summating potential (SP)}
Observé pour la première fois par Davis en 1950, chez le cobaye, c’est un signal électrique complexe et multi composant. Il précède la réponse nerveuse et correspondrait à une polarisation particulière des cellules ciliées de l’organe de Corti.
D’après les travaux de Whitfield et Ross (1965)(17), ce type de polarisation se produit lors des déplacements non linéaires de la membrane basilaire, provoqués par une stimulation sonore de haute intensité. C’est pourquoi le SP n’est présent, sur le tracé, que lors des stimulations sonores suffisamment intenses (> 70 dB)(fig. 2).
L’amplitude de SP dépend, donc, de l’intensité et de la fréquence du stimulus sonore et elle est directement proportionnelle à la somme des déplacements de la membrane basilaire. Il n’est pas sensible à la polarité du stimulus, et ne s’élimine pas lors de l’inversion de phase.
Réponses nerveuses :
Après l’étape sensorielle de la réception de la vibration sonore par les cellules ciliées, a lieu l’étape ultime de la transformation du signal sonore en signal nerveux.
Le signal sonore est transformé en un potentiel d’action unitaire par la stimulation du fibre nerveuse connectée à la cellule ciliée. Ces potentiels unitaires cheminent dans les fibres nerveuses (30.000 fibres) et forment un potentiel d’action global ou composite au niveau du nerf auditif.
L’observation des potentiels d’action unitaires est très délicate (utilisation de micro-électrode) et ne donne qu’une information limitée sur le fonctionnement de l’ensemble de la cochlée et du nerf auditif. Par contre, l’observation des phénomènes globaux est à la fois plus facile (utilisation de macro-électrode) et permet d’obtenir une information globale sur l’ensemble des structures cochléaires.
L’EcoG, comme pour les réponses sensorielles, permet d’observer le résultat global de cette étape de transformation.
· Le potentiel d’action global du nerf auditif : {Action potential (AP)}
Observé pour la première fois par Saul et Davis, en 1932, au sein des réponses cochléaires, c’est la combinaison des potentiels d’action unitaires.
Sur l’EcoG, l’AP d’amplitude plus élevée que celle du SP, apparaît juste après ce dernier et forme la composante principale du complexe de l’onde I.
L’observation de ce phénomène global nécessite une excitation simultanée de l’ensemble des fibres nerveuses. Si les fibres répondent à des instants différents sans aucun synchronisme, le phénomène global, représentant la somme de ses réponses, sera statistiquement nul. Le stimulus doit être, pour pouvoir stimuler simultanément l’ensemble des cellules ciliées, le plus bref possible (des transitoires). Le stimulus le plus apte à pouvoir provoquer ce potentiel d’action est le clic sonore.
Principes techniques de L’EcoG :
La réalisation de l’EcoG nécessite un dispositif électronique permettant d’une part la stimulation de l’oreille testée, et d’autre part le recueil des signaux électriques. Les réponses recueillies subissent, par ce dispositif, une sommation algébrique dont le but est d’éliminer le bruit de font et d’extraire, sur une courbe moyennée, les seules réponses électriques provoquées par la stimulation sonore au niveau de la cochlée. Le procédé employé est identique à celui utilisé pour la réalisation des PEA, et sa description n’entre pas dans le cadre de ce mémoire.
L’EcoG se réalise dans un endroit calme, silencieux et électriquement isolé. Le sujet examiné doit être au repos, allongé, les yeux fermés et le plus détendu possible.
La stimulation sonore est transmise à l’oreille interrogée par l’intermédiaire d’un casque classique.
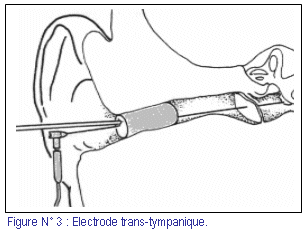
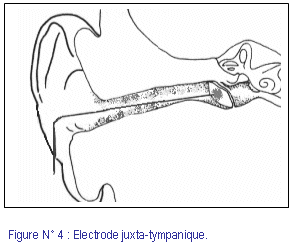
Le recueil se fait grâce aux trois électrodes (positive, négative et neutre), dont une électrode (de polarité négative) doit être posée le plus proche possible de la cochlée. Selon l’emplacement de cette électrode, trois méthodes ont été décrites : (fig. 3, 4)
1) EcoG trans-tympanique : l’électrode est posée sur le promontoire.
2) EcoG juxta-tympanique : l’électrode est posée sur le tympan en regard de la fenêtre ronde.
3) EcoG trans-méatique : l’électrode est posée dans le conduit auditif externe.
Choix de l’emplacement de l’électrode :
Les premières EcoGs, chez l’homme, (Lempert et coll. 1947), avaient été réalisées en positionnant une électrode sur la membrane de la fenêtre ronde. Des études chez l’homme et chez l’animal, [Arts et coll., Krueger et coll., Bar el dine et coll. (3, 7, 25)] ont ensuite prouvé que c’était l’emplacement permettant l’enregistrement des réponses cochléaires les plus importantes en amplitude. Mais, la nécessité d’une intervention chirurgicale, limitant cette méthode, a conduit à retrouver des emplacements de moins en moins invasifs.
· EcoG trans-tympanique :
La méthode trans-tympanique (fig. 2 , 3), qui a été utilisée depuis 1967 [Aran et coll., Gibson (2, 17)], propose de poser une électrode métallique rigide et fine sur le promontoire, en transperçant le tympan. La plupart des études cliniques réalisées, jusqu’à ce jour, utilisent cette méthode. Le résultat obtenu est de grande qualité, mais son inconvénient majeur est son caractère invasif vis à vis du tympan. Sa réalisation nécessite, en effet, une anesthésie générale, surtout chez l’enfant.
· EcoG trans-méatique :
L’emplacement trans-méatique a été proposé par quelques équipes. Il consiste à poser une électrode cylindrique dans le conduit auditif externe. Roland et coll. (42), ainsi que Schwaber et coll. (47) ont réalisé des études cliniques dans lesquelles ils comparent ces deux méthodes (trans-tympanique et trans-méatique). Bien que, cette technique soit beaucoup plus simple à réaliser, les signaux électriques obtenus sont de faible amplitude. Les auteurs notent, de plus, une grande variabilité inter-individuelle et une reproductibilité inconstante.
· EcoG juxta-tympanique :
L’emplacement juxta-tympanique (fig. 4) qui propose de poser l’électrode au contact du tympan, a été utilisée pour la première fois en 1972 par Cullen et coll. [Ruth et coll. (43)]. Ensuite, cette méthode a été reprise et modifiée en 1987 par Stypulkowski et Staller [Ruth et col., Laureano et coll. (43, 28)]. De réalisation plus simple sans recours à l’anesthésie générale, cette méthode permet d’obtenir des électrocochléogrammes de très bonne qualité. Les réponses cochléaires y sont d’amplitude suffisante, et reproductibles. Les équipes qui ont utilisé cette méthode [Campbell et coll., Pou et coll. (11, 40)], sont peu nombreuses.
Les avantages techniques de l’EcoG juxta-tympanique, nous ont conduit à mettre au point un protocole qui permet de réaliser des EcoGs de façon plus systématique, et même en ambulatoire. Ce protocole, utilisé pour l’étude clinique de ce mémoire, sera décrit dans le 5e chapitre.
Choix de la stimulation sonore :
L’essentiel pour provoquer une réponse globale du nerf auditif, facilement observable, est de déclencher simultanément les fibres nerveuses. Pour cela, le stimulus doit être de durée brève. La stimulation la mieux adaptée et la plus utilisée est le clic sonore [Aran et coll., Arts et coll., Gibson, Mori et coll. (2, 3, 17, 34)].
· Clic sonore :
Il correspond à une impulsion rectangulaire d’une durée égale à 100 µs. Le clic est l’équivalent du choc électrique utilisé en neurophysiologie pour stimuler directement un nerf. Non filtré, son spectre de fréquence est continu et étendu (de 1000 à 4000 Hz). C’est dans le domaine des sons brefs, l’équivalent du bruit blanc par rapport aux sons purs dans le domaine des sons permanents. Le spectre large du clic est un avantage, car ainsi une grande partie de la cochlée est simultanément stimulée. Cependant, les informations recueillies en réponse à un clic, et en particulier le seuil, ne permettront pas de préciser la sensibilité en fréquence de la cochlée.
Le clic non filtré peut, donc, être utilisé en une série de clics en condensation ou en raréfaction. L’alternance de phase (clics alternés) permet l’élimination des potentiels microphoniques (fig. 1).
· Autres stimuli sonores :
Le clic non filtré n’est pas le seul moyen d’évoquer une réponse du nerf auditif. Le clic filtré ou « Tone pip » [Aran et coll., Koyuncu et coll. (2, 24)] est un clic dont la bande de fréquence est étroite. En filtrant l’impulsion rectangulaire, servant à produire le clic, dans un filtre passe-bande, on obtient un clic dont le spectre fréquentiel est centré sur une fréquence donnée. En générale, les fréquences utilisées sont : 1000 Hz et 4000 Hz.
Les bouffées tonales ou « Tone burst » sont des tranches de son pur dont l’établissement peut être plus ou moins rapide et la durée plus ou moins longue. Elles sont filtrées et utilisées, en générale, à 1000 ou 2000 Hz. Pour Koyuncu et coll. (24) la stimulation en bouffée tonale augmente la sensibilité de l’EcoG. Dans leur étude, ils apportent que 4/40 patients (10%) avaient une EcoG normale en stimulation par clic. Ces mêmes patients ont eu une EcoG anormale lors de stimulation par bouffée tonale. Orchik et col. (38) notent une meilleure interprétation, et une possibilité d’étudier dans les fréquences graves, lors d’utilisation des bouffées tonales.
Dans notre expérience, la stimulation par bouffées tonales ne permet pas d’obtenir la même reproductibilité des tracés que par stimulation avec clic sonore.
Tracé de l’EcoG : (stimulation par clic alterné non filtré) :
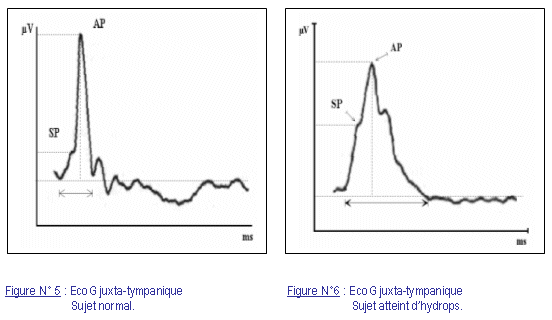
La courbe de l’EcoG est caractérisée par un complexe, survenant après l’artéfact de stimulation, qui correspond au complexe I des PEA. Ce complexe est représenté dans notre étude en polarité positive par similarité aux PEA (fig. 2, 5, 6).
Il se décompose, dans les hautes intensités de stimulation, en deux parties. La première partie du complexe est de faible amplitude, formant l’épaule du complexe I, elle correspond à une polarisation particulière des cellules ciliées (SP). Ensuite la déflexion atteint son point culminant et retourne à la ligne de base. Cette deuxième partie, d’amplitude plus élevée, correspond aux potentiels électriques produits par l’ensemble des fibres nerveuses stimulées (AP). Dans certains cas pathologiques, la déflexion de la repolarisation est confondue dans le complexe I, formant une deuxième épaule quasi symétrique au SP. Ce qui donne un aspect élargi, caractéristique, au complexe I.
Interprétation du tracé de l’EcoG : (dans le diagnostic de l’hydrops)
L’interprétation de l’EcoG est basée sur le rapport d’amplitude du SP et de l’AP. L’aspect morphologique du complexe I peut aussi être pris en considération [Koyuncu et coll. (23, 24)]. Un complexe normal est d’aspect étroit et la déflexion de la repolarisation ne fait pas totalement partie du complexe I (fig. 5, 6).
Il existe une grande variabilité inter-individuelle et les facteurs influençant la qualité de l’EcoG sont [Roland et coll. (41)] :
- L’emplacement de l’électrode.
- La stimulation sonore (intensité, fréquence).
- Le seuil auditif du sujet examiné.
- Les conditions de l’examen.
Pour le rapport d’amplitude de SP/AP, les auteurs rapportent des normes plus ou moins uniformes.
è EcoG trans-tympanique avec stimulation en clics alternés :
De façon quasi unanime, ce rapport est considéré normal s’il est inférieur à 0,25 pour les intensité supérieure à 70 dB. Pour Gibson (18) il existe une relation directe entre la valeur du SP/AP et l’intensité de la stimulation. Les normes proposées par Gibson, en fonction de l’intensité du stimulus sont :
· de 0 à 40 dB : SP/AP < 0,50
· de 40 à 70 dB : SP/AP < 0,35
· plus de 70 dB : SP/AP < 0,25
A titre indicatif, d’autres valeurs retenues comme normales par d’autres auteurs :
· Filipo et col. (14) : SP/AP < 0,27
· Arts et col. (3) : SP/AP < 0,30
· Orchik et col. (35, 37) : SP/AP < 0,40
è EcoG trans-tympanique avec stimulation en bouffée tonale :
D’après Gibson (18) l’amplitude absolue normale du SP doit être inférieure à 3 µV en stimulation par bouffées tonales de 1000 Hz.
è EcoG juxta-tympanique avec stimulation en clics alternés :
La valeur du SP/AP est considérée normale, dans notre étude, si elle est inférieure à 0,35. Les auteurs utilisant cette méthode retiennent soit la valeur de 0,30 [Campbell et coll. (11)] ; soit, comme nous, la valeur de 0,35 [Pou et coll. (40)].
è EcoG trans-méatique avec stimulation en clics alternés :
Mori et coll. (34), dans une étude réalisée en méthode trans-méatique et stimulation à 80 dB, considèrent le rapport SP/AP normal s’il est inférieur à 0,43.
APPLICATIONS CLINIQUES DE L’EcoG :
L’EcoG est essentiellement utilisée dans 3 buts :
1. Mise en évidence d’une onde I lorsqu’elle n’apparaît pas sur les tracés des PEAp, et détermination du seuil auditif.
2. Recherche d’une activité cochléaire résiduelle chez l’enfant, dans le cadre du bilan de l’implant cochléaire.
3. Diagnostic d’un trouble de pression endolymphatique (hydrops endolymphatique ; fistule périlymphatique) [Badr el dine et coll., Gibson, Meyerhoff et coll., Uchida et coll. (7, 17, 32, 51)].
Détermination du seuil auditif :
L’application de l’EcoG, en complément des PEA, à la recherche d’un seuil auditif chez l’adulte [Gibson, Laureano et coll., Ohashi et coll. (17, 28, 36)] ou chez l’enfant sourd profond [Aso et coll., Gibson, Wong et coll. (4, 17, 53)], est bien connue et n’a pas besoin d’être justifiée. En effet, la sensibilité de l’EcoG pour détecter une activité cochléaire est plus grande que les PEA. Lorsqu’une courbe de PEA est plate, l’EcoG peut mettre en évidence la présence d’un reste auditif.
Monitoring peropératoire :
L’EcoG a été utilisée dans le monitoring peropératoire notamment dans la chirurgie du neurinome de l’acoustique [Sandra et coll. (44)]. L’étude des latences des AP et SP pendant l’intervention, peut remplacer ou compléter le monitoring par les PEA et apporter au chirurgien une sécurité supplémentaire.
Surdités de perception :
Le rapport d’amplitude de SP/AP n’est pas modifiée dans les surdités dont l’étiologie n’est pas un trouble de pression endolymphatique [Liu et coll. (30)]. Par contre, elle permet, parfois, de retrouver l’origine hydropsique de certaines surdités brusques [Filipo et coll. (15)]. La maladie de Ménière pouvant se déclarer par un épisode de surdité brusque, il est clair que l’EcoG peut apporter un élément diagnostique avant que le tableau clinique ne soit complet.
Troubles de pression endolymphatique :
Tout trouble de pression endolymphatique, hypopression ou hyperpression, se traduit sur la courbe de l’EcoG par une augmentation de l’amplitude du SP [Badr el dine et coll., Gibson, Meyerhoff et coll., Uchida et coll. (7, 17, 32, 51)]. L’amplitude de l’AP restant inchangée ou voire diminuée, le rapport du SP/AP augmente.
Nous avons vu que le SP était le résultat d’une polarisation particulière des cellules ciliées induite par les déplacements non linéaire de la membrane basilaire, en réponse à un stimulus sonore de haute intensité [Aran et coll., Gibson (2, 17)]. Les déformations des structures des compartiments endolymphatiques (la membrane basilaire, la membrane de Reissner), induites par le déséquilibre pressionnel qui règne dans les milieux endolymphatiques, entraînent une perte d’accouplement entre les cellules ciliées et la membrane tectoriale. Ce phénomène serait responsable d’une part de la surdité (dans les premiers stades de l’hydrops) et d’autre part de l’augmentation d’amplitude du SP [Ge et coll. (16)].
Le rapport SP/AP est donc augmenté dans l’hydrops endolymphatique (secondaire ou idiopathique), dans les fistules périlymphatiques, et tout changement biochimique de la composition de l’endolymphe perturbant l’équilibre pressionnel des compartiments endolymphatiques [Uchida et coll. (51)].
Modèle animal :
Les expérimentations chez l’animal (cochon d’Inde) confirment l’augmentation de l’amplitude du SP dans l’hydrops endolymphatique induit.
Badr el dine et coll., Uchida et coll. (8, 51) ont réalisé une série d’EcoG chez les cochons d’Inde avant et après l’oblitération du sac endolymphatique. L’étude histopathologique confirmait le développement de l’hydrops en objectivant une déformation de la membrane de Reissner. L’analyse des courbes obtenues met en évidence une augmentation de l’amplitude du SP, de façon significative, dans les premiers stades de la formation de l’hydrops. L’amplitude du SP redevient à la normale, lorsque la membrane de Reissner est très distendue. Ces constatations chez animal sont aussi valables chez l’homme, mais l’étape ultime de l’hydrops expérimental n’est peut être jamais observée chez l’homme.
EcoG et la maladie de Ménière :
L’apport diagnostique de l’EcoG dans la maladie de Ménière est indiqué par de multiples études. L’augmentation du rapport SP/AP est très souvent observée chez les patients atteints de la maladie de Ménière, quel que soit le stade de la maladie. Il ne semble pas exister de rapport direct entre l’importance de cette modification et le grade ou l’ancienneté de la maladie [Dornhoffer et coll., Levine et coll. (14, 29)].
La réalisation d’EcoG après l’injection ou l’ingestion des produits osmotiques (comme nous avons vu dans le 3e chapitre), montre une tendance à la normalisation du rapport du SP/AP chez certains patients. Cette influence des produits osmotiques est le témoin de la réversibilité de l’hydrops chez ces patients. Ce phénomène n’est pas observé avec d’autres traitements de type : chirurgie du sac endolymphatique, injection d’aminosides ou de corticoïdes [Orchik et coll. (37)].
Les facteurs intrinsèques influençant la positivité de l’EcoG [Koyuncu et coll., Pou et coll. (23, 40)] sont :
- L’importance de la surdité.
- Le type de la surdité.
- L’ancienneté de la maladie.
D’après Pou et coll. (40), l’EcoG a plus de chance d’être positive si :
- La surdité est fluctuante.
- La surdité est peu importante (< 40 dB).
- La maladie évolue depuis moins de 48 mois.
Intérêts de l’EcoG dans la maladie de Ménière :
Les usages potentiels de l’EcoG dans la maladie de Ménière [Levine et coll. (29)] sont *:
a) Elle permet un diagnostic précoce de la maladie**.
b) Sa normalisation sous l’influence des produits osmotiques (plus sensible que les critères audiométriques) est le témoin de la réversibilité de la symptomatologie, guidant le clinicien dans sa démarche thérapeutique.
c) Elle peut être utilisée dans la surveillance clinique chez les patients traités par des produits osmotiques à long terme.
Spécificité et la sensibilité de l’EcoG :
Nous avons décrit, ci-dessus, les valeurs considérées comme normales pour le rapport du SP/AP. Certains auteurs, en gardant une marge, considèrent que l’EcoG est seulement douteuse si la valeur du SP/AP est comprise entre 0,35 et 0,50 [Arts et coll., Pou et coll. (3, 40)]. Cette considération conduit à diminuer la sensibilité de l’EcoG.
La sensibilité est définie comme le nombre de tests positifs chez les sujets atteints, et la spécificité est le nombre de tests négatifs chez les sujets non atteints.
La plupart des études retrouvent une spécificité aux alentours de 95%. Tandis que la valeur de la sensibilité varie du 57% à 77%, selon la norme du SP/AP définie par l’étude. Pour Gibson (18) la spécificité de l’EcoG est de 94% et sa sensibilité est de 77% (en méthode trans-tympanique, SP/AP > 0,25). Pou et coll. (40) trouvent la même spécificité à 1% près, mais la sensibilité retrouvée est de 57% (méthode juxta-tympanique, SP/AP > 0,50). Sass et coll. (45), rapportent une spécificité de 95% et une sensibilité de 62% (méthode trans-tympanique en bouffées tonales). Ge et coll. (16), en méthode trans-tympanique et en prenant la valeur de 0,40 pour SP/AP, retrouvent une spécificité égale à 81,3%.
Dans notre expérience, nous avons retenu la valeur de 0,35 comme la limite de la normalité du SP/AP. Toute valeur supérieure à 0,35 est considérée indicative d’hydrops endolymphatique. En plus de la mesure de cette valeur, l’aspect morphologique du complexe I est, aussi, pris en considération. Dans l’hydrops, le complexe est souvent élargi. Cet aspect caractéristique, correspond au fait que la déflexion qui suit normalement le complexe est plus ou moins comprise dans le complexe. Du fait de l’augmentation de l’amplitude du SP, elle devient plus ample et apparaît quasi symétriquement au SP dans la partie descendante de l’AP.
Au total, une EcoG positive est un argument objectif pour le diagnostic de l’hydrops endolymphatique (spécificité = 95%), mais une EcoG négative n’élimine pas ce diagnostic (sensibilité= 77%).